Il y a peu, il était admis que les collemboles ne maitrisaient pas leurs sauts au point d'en faire un moyen de locomotion ordinaire. Ces sauts, issus d’une détente explosive de leur furca, étaient alors considérés comme un réflexe de fuite assorti de trajectoires aléatoires. Cependant, l'article consacré au collophore, montre que cet organe peut être utilisé comme moyen de fixation au substrat, lors de la réception d’un saut. On peut donc considérer que le collembole dispose, au moins partiellement, d’une capacité à contrôler certaines phases de son vol.
La dynamique de la locomotion animale suscite un intérêt croissant chez les biologistes, ingénieurs et mathématiciens qui y voient des potentiels novateurs, aussi bien dans des secteurs comme la robotique que ceux des dispositifs médicaux (prothèses). On sait que la locomotion des arthropodes représente une de leur plus forte dépense d’énergie. En effet, le bilan énergétique de leur locomotion se dégrade d'autant que leur masse corporelle décroit. Au cours de leur évolution, les collemboles, dont le poids est très faible, ont donc probablement été conduits à adopter des modèles susceptibles d'améliorer le ratio dépense/efficacité de leur système locomoteur. La capacité à fuir le danger étant, de surcroît, une de leur principale condition de survie.
Pour approcher les mécanismes en jeu durant le saut, on doit prendre en compte plusieurs composantes. Par exemple, la nature de l'impulsion initiée par la furca ou les mouvements et postures du corps du collembole durant le vol. Les prises de vue réalisées sur des sauts, ainsi que les études morphologiques approfondies, apportent de nouveaux éléments qui n’étaient pas accessibles à l’observation conventionnelle, ne serait-ce qu’en raison de la brièveté des événements concernés, quantifiables en ms. (millièmes de secondes) ou de la complexité morphologique de l’appareil sauteur. On est désormais capable de mesurer vitesses et accélérations au cours des vols et de prendre en compte, grâce à la modélisation informatique, des paramètres d’aérodynamisme (effets de l’air sur les corps en mouvement) ou d’hydrodynamisme (effets des liquides incompressibles sur les corps en mouvement).
J’évoque ici, "sans équation", l'approche des scientifiques sur la cinématique des sauts de certains collemboles. Ce chapitre m’a été inspirée par "Décollage directionnel, redressement aérien et atterrissage en adhérence de collemboles semi-aquatiques" (nov. 2022) [1], publication d’une équipe de chercheurs de l’université du Maine (USA). J’ai contacté un des co-auteurs, Victor Ortega Gimenez, qui m’a fait parvenir des vidéos dont il m’a autorisé l’utilisation. Pour les besoins de la transcription ecrite, une partie des illustrations qui suivent sont donc des copies d’images sélectionnées à partir de séquences vidéo. Enfin une partie des schémas qui suivent sont des adaptations réalisées par mes soins, car l’étude précitée est protégée par copyrights.
Éléments de morphologies :
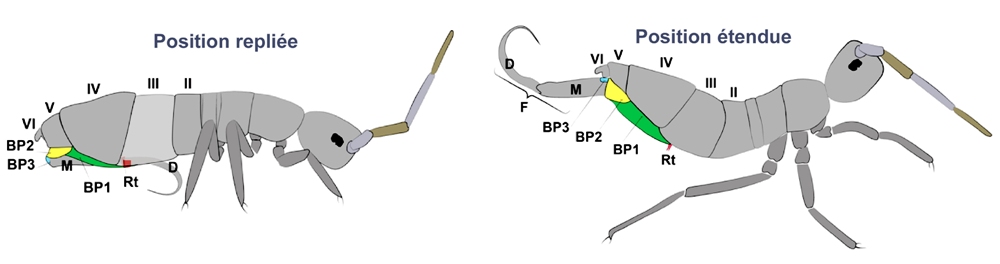
En complément de l'article "Tube ventral structure fine", j’évoque ici quelques éléménts d'anatomie déterminants lors des sauts. Une partie des illustrations qui suivent est issue d’un article écrit par Fabio Goncalves de Lima Oliveira [2] : « Sur les collemboles : une étude morpho-fonctionnelle de l’appareil sauteur ». L’observation à l’œil nu ne permet pas d’appréhender la complexité de l’appareil sauteur du collembole, longtemps considéré comme une simple catapulte. Ci-dessous, deux positions de la furca (F) d’un Entomobryomorphe. Les tiges de la sclérite basales (BP1, 2, 3) y sont représentées en couleur.
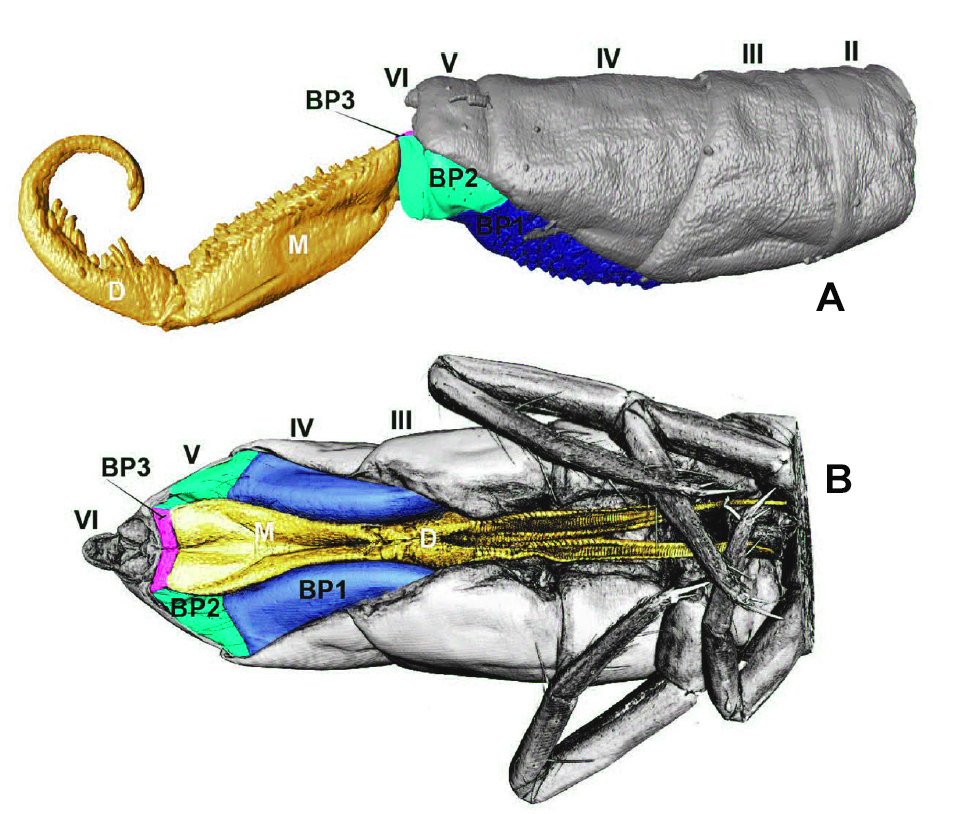
Ci-dessous, reconstitution MicroCT révélant les plaques basales d’Orchesella cincta. La figure « A » est une vue latérale, depuis le second segment abdominal jusqu’à l’extrémité de la furca. Elle correspond au schéma précédent « position étendue ». La figure « B » est une vue ventrale de l’appareil sauteur avec la furca en position de repli, fixée par le rétinacle.
Les scientifiques qui s’interrogent sur la nature des mécanismes qui provoquent le saut émettent deux hypothèses :
1) L’énergie élastique (résultant d’une action musculaire) nécessaire au mécanisme du ressort serait directement stockée dans la sclérite basale du quatrième segment abdominal.
2) Le saut est déclenché par un bref accroissement de la pression hémolymphatique.
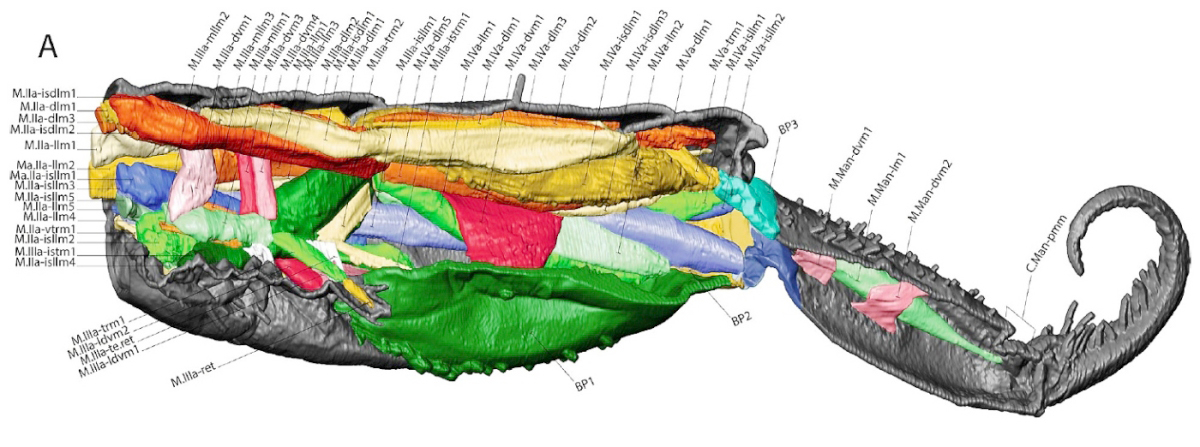
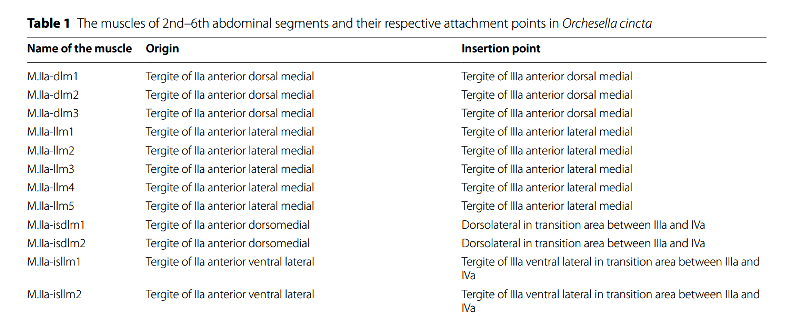
Les études réalisées à ce jour laissent penser que la réponse pourrait être une combinaison des deux : « système musculaire + pression hydraulique ». Afin de comprendre ces aspects morpho-fonctionnels, les chercheurs ont approfondi l’étude anatomique de l’appareil sauteur, comme en témoigne, ci-aprés, la reconstitution morphologique microCT* en vue latérale interne des segments abdominaux 2 à 6. On y distingue les muscles et la cuticule de l’appareil sauteur d’un Orchesella cincta. Chaque muscle est répertorié et classé dans une nomenclature qui définit son nom et les points de rattachement de ses deux extrémités (voir extrait sous l’illustration).
* Remarque : Le niveau de détail, indispensable pour aborder les mécanismes de la détente musculaire, est quasiment inaccessible par les méthodes conventionnelles, basées sur l’observation après dissection. Il n'est accessible qu’en ayant recours à la technologie nommée "micro-CT" ou micro-tomodensitométrie. Cette reconstitution en imagerie 3D (tridimensionnelle) utilise les rayons X pour voir l'intérieur d'un sujet. Elle génère des images détaillées de la structure interne des échantillons de petites dimensions et autorise des résolutions de 100 nanomètres, soit 0,001 mm. Cette précision microscopique est obtenue avec un montage comme celui décrit ci-dessous.
Méthodologie :
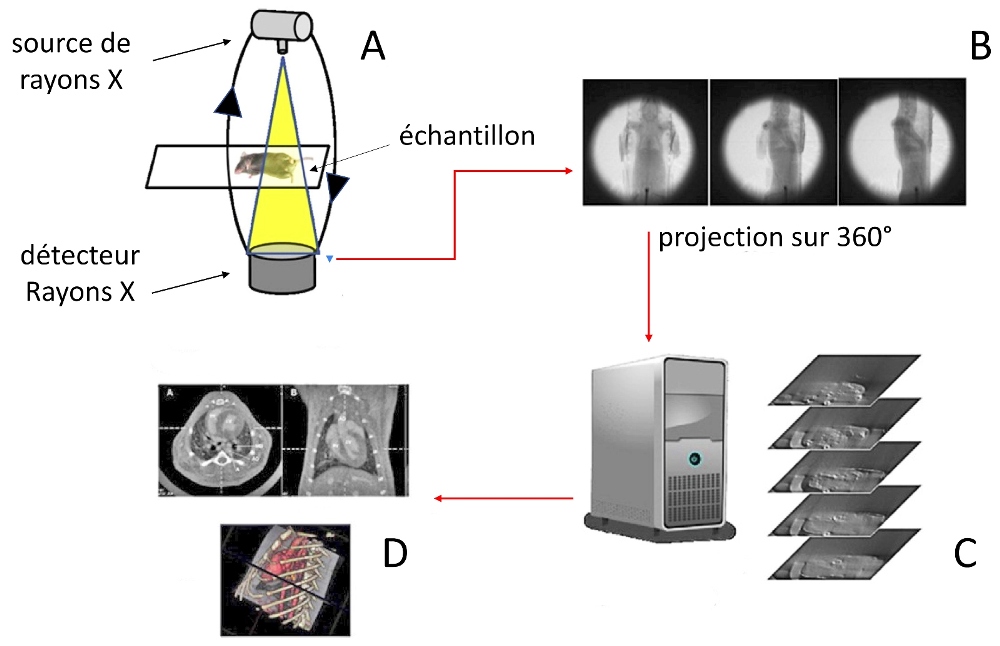 - L’échantillon à étudier est exposé à une source de rayon X (A).
- L’échantillon à étudier est exposé à une source de rayon X (A).
- Un balayage angulaire (par rotation) est effectué autour de ce sujet pour prendre, tranche par tranche, plusieurs centaines de clichés bidimensionnels (B).
- Un prétraitement informatique (C) de ces clichés (étalonnage photométrique) les transforme en logarithmes. Processus également nommé : Transformées en rayon X à faisceau conique des coefficients d’atténuation linéaire.
- Ces logarithmes sont traités par le calcul, à l’aide d’un algorithme nommé algorithme de FeldKamp. Ce dernier intègre les paramètres géométriques du balayage afin de reconstituer des images tomographiques dépourvues de tout artefact qui pourrait être issu, par exemple, d’un défaut d’alignement.
Les images ainsi recomposées et colorisées peuvent être exploitées. Les diverses mesures dimensionnelles effectuées à partir de ces dernières sont un reflet fiable du sujet. Cette technologie révèle, au niveau le plus fin, la morphologie de l’ensemble de l’appareil sauteur et permet de modéliser les différents leviers qui interagissent en son sein, comme le montre la figure suivante qui montre la modélisation d'une furca en position dépliée.
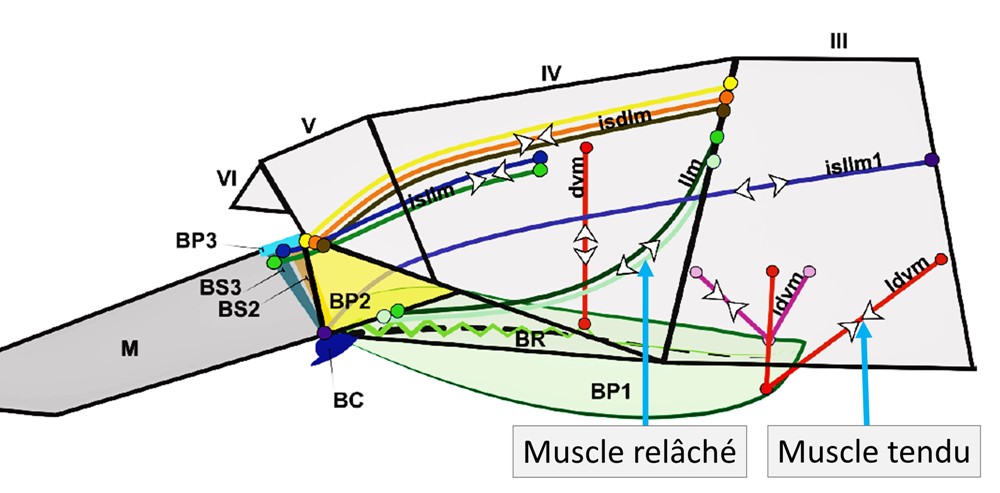
On note que chaque muscle, ou faisceau de muscles, est assorti de paires de flêches qui, selon leur orientation, indiquent si ce dernier est tendu ou relâché. Cette même modélisation, établie pour d’autres positions de la furca, indique que les stratégies et mécanismes impliqués lors des sauts, varient entre différentes espèces possédant des segmentations distinctes de forme cylindrique (Tomocerus.sp et Orchesella.sp) et celle de forme globulaires (Sminthurus.sp, Bourletiella.sp). Le rôle essentiel des plaques basales (BP.) est, par ailleurs, identifié en tant que siège du stockage de l’énergie libérée sous forme d’énergie cinétique au moment du saut. La fonction de déclencheur du rétinacle, en plus de celle déjà connue de verrouillage de la furca, est également confirmée.
Méthodologie de l’étude du saut :
La modélisation du système sauteur est un élément essentiel à la compréhension de son fonctionnement. Cependant, elle ne saurait s’affranchir d’une analyse cinématique. En effet, les différentes phases d’un saut : décollage, vol et atterrissage, outre de mettre en œuvre la musculature décrite ci-dessus, font appel à d’autres moyens de « pilotage » auxquels à recours le collembole. Ci-aprés, schéma de la méthodologie mise en place pour l’analyse du vol d’un Isotomurus retardatus qui possède un corps allongé et segmenté, comparable à celui d’Orchesella cincta.
Description : Un échantillonnage d’Isotomurus retardatus est placé sur un récipient de faible profondeur recouvert d’eau (reconstitution en laboratoire du milieu dans lequel évolue ce collembole). Cette zone aquatique est éclairée par une lampe à halogénures métalliques produisant une intense lumière blanche (neutre), indispensable pour éclairer les sujets durant les prises de vue à cadence rapide.
 Les sauts des collemboles sont filmés à une cadence de 1000 à 10.000 im/sec. Pour les plans rapprochés, on utilise un objectif Canon MP-E65 mm (voir article "Materiel photographique") et un objectif Canon EF 200 mm pour les plans larges. La faible taille des collemboles, la brièveté de leurs vols et la limitation des distances focales des montages optiques retenus génèrent beaucoup de déchets de prise de vue.
Les sauts des collemboles sont filmés à une cadence de 1000 à 10.000 im/sec. Pour les plans rapprochés, on utilise un objectif Canon MP-E65 mm (voir article "Materiel photographique") et un objectif Canon EF 200 mm pour les plans larges. La faible taille des collemboles, la brièveté de leurs vols et la limitation des distances focales des montages optiques retenus génèrent beaucoup de déchets de prise de vue.
Un long travail de visualisation préalable est nécessaire pour en extraire des séquences exploitables. Parallèlement à l’aspect technique, lié à la réalisation des films, il faut aussi s’assurer de pouvoir effectuer certaines mesures, tout au long des phases de vol, comme la vitesse instantanée υi ou la position angulaire du corps ωi du sujet.
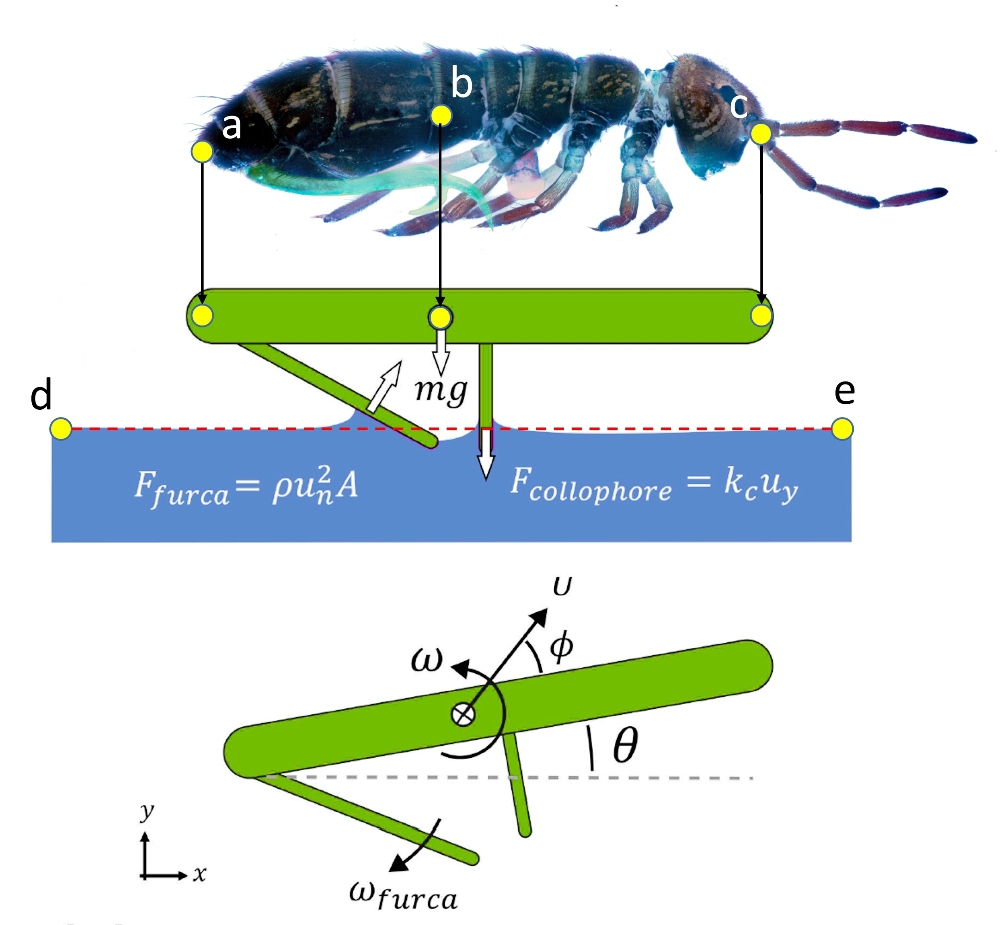 À ces fins, les chercheurs ont mis en place des repères virtuels sur le corps modélisé du collembole (illustration ci-contre).
À ces fins, les chercheurs ont mis en place des repères virtuels sur le corps modélisé du collembole (illustration ci-contre).
La numérisation de trois points déterminants (ici en jaune) du collembole, permet de recueillir des données tout au long de sa trajectoire selon un repère à deux dimensions, relativement au point central (b), considéré comme centre de gravité. Les deux autres repères, (a) et (c), respectivement situés à l’extrémité de l’abdomen et à la pointe de la tête, permettent, quant à eux, de modéliser les mouvements propres du collembole qui évolue sur cette trajectoire.
Enfin, deux points distants sur la surface de l’eau, (d) et (e), établissent le référentiel bidimensionnel (x,y). Ainsi, grâce à ces repères, on connait, à chaque instant, la posture corporelle, l’angle de décollage θ et la hauteur par rapport à l’eau. De ces valeurs on déduit, par le calcul, les paramètres instantanés du vol, comme vitesse, accélération, rotations, etc… Ces calculs étant réalisés sur base des équations classiques de la cinématique ou de l’hydrodynamique.
L'étude qui permet aux chercheurs d’analyser différentes phases de la trajectoire du collembole semi-aquatique Isotomurus retardatus a montré que cette espèce est capable d’effectuer des sauts directionnels, des redressement aériens rapides et des réceptions, à la surface de l’eau, parfaitement maîtrisés. Ci-dessous, huit vues d'une séquence de décollage sur lesquelles la furca est colorisée en jaune, le collophore en rouge et la gouttelette d’eau en bleu.
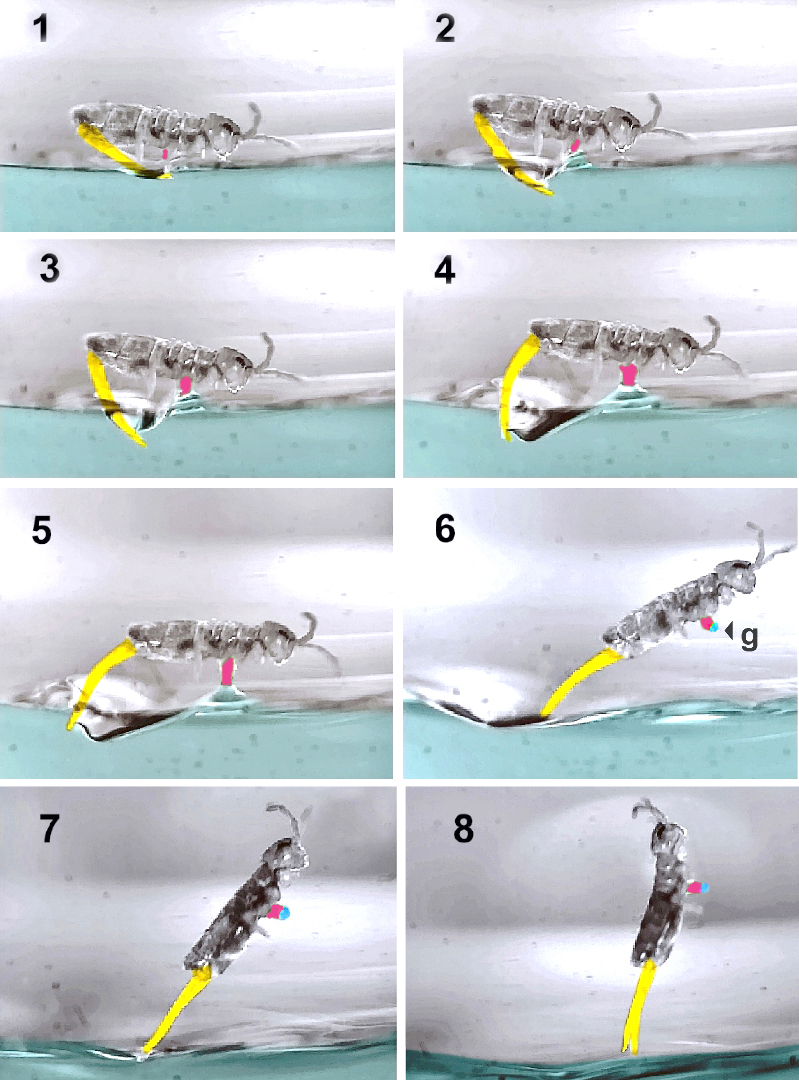
Description :
- Avant de sauter, le collembole s’ancre à la surface de l’eau à l’aide du collophore (1).
- Au début du déploiement de la furca, ce dernier reste en contact avec l’eau (2, 3, 4, 5) et la pointe de son abdomen est maintenue la plus éloignée possible de la surface.
- Au décollage (6, 7, 8), le collophore emprisonne une gouttelette (g) d’eau qui pèse approximativement 3% de la masse corporelle et qui joue le rôle de contrepoids, pour aider au redressement aérien.
Le temps écoulé entre chaque image est d’environ 0.4 ms, ce qui donne une idée du laps séparant le début de la détente et le moment où la furca n’est plus en contact avec l’eau, soit un total de 3,2 ms.
Ci-dessous, modélisation d’un décollage, qui présente chaque position angulaire de la furca (manubrium = rouge , dente + mucron = vert) avec des intervalles de 0.2 ms. Les très courtes durées mesurées durant ces événements révèlent des valeurs impressionnantes : Par exemple, la vitesse de déplacement constatée atteint jusqu’à 280 fois la longueur du corps par seconde. Isotomurus mesurant environ 2,3 mm, on en déduit une vitesse lors du décollage de 0,64 m/s, ce qui, ramené à l’échelle d’un homme de 1.75m [1], correspond à une vitesse supersonique de 1715 km/h (Mach 1.4). Les accélérations mesurées se situent quant à elles autour de 50 ± 30 m/s² soit, entre 2 et 5 G. À titre de comparaison, 2 G est l’accélération maximale d’une formule un et 5G correspond au seuil de perte de la vision centrale (voile noir) d’un pilote d’avion de chasse.
Les calculs effectués à partir de ces modèles montrent que, durant la phase initiale du décollage, la force d’adhérence à l’eau du collophore entre, pour une bonne part, dans le contrôle directionnel de l’impulsion. Cette force d’adhérence est due à la microstructure du collophore et à la tension superficielle des fluides (propriété qui permet aux collemboles de demeurer sur ses pattes à la surface de l’eau sans couler, ou, à d’autres animaux, comme les Gerris, de se déplacer à la surface des mares).
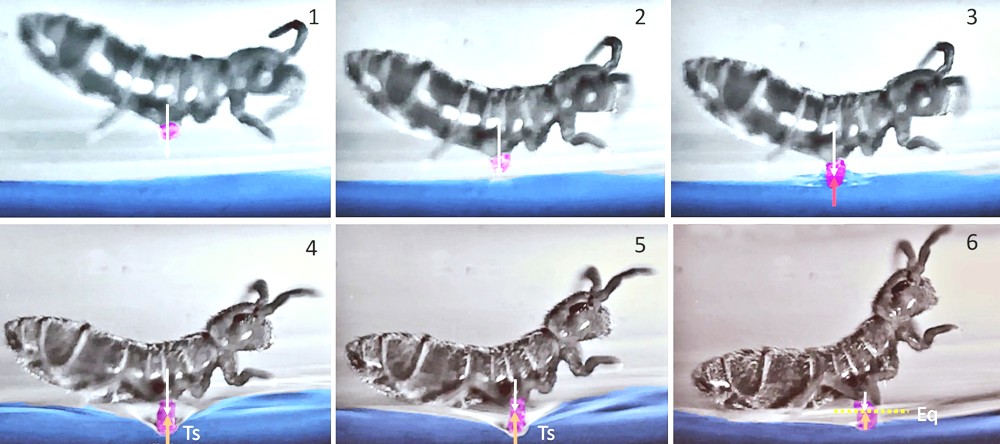
Les atterrissages, également modélisés, ont permis d’établir l’efficacité du collophore comme stabilisateur à la réception. Page suivante, une succession d’images montre l’usage du collophore (colorisé en rouge) lors d’une réception.
À l’issue d’un saut, le collembole arrive à la surface de l’eau (1, 2) avec des vitesses d’impact moyennes de 0.5 m/s. En raison de la forme arquée qu’il adopté durant son vol, le contact avec le liquide s'effectue par l’intermédiaire du collophore (3). Il s’en suit un passage au dessous du niveau de l’eau, jusqu’à une profondeur équivalente à 60% de la largeur abdominale, soit 0.2 mm (4, 5).
Cependant, la tension superficielle a pour effet une déformation incurvée de la surface sans que le collembole ne soit immergé. Ces mêmes forces exercent une contre-poussé (Ts) au niveau de la zone de contact (colophore/eau) qui permet d’arriver au point d’équilibre (6). Le collembole est alors stabilisé. Entre l’instant ou le collophore entre en contact avec l’eau et celui où la stabilisation est achevée, il s’est ici écoulé environ 4 ms.
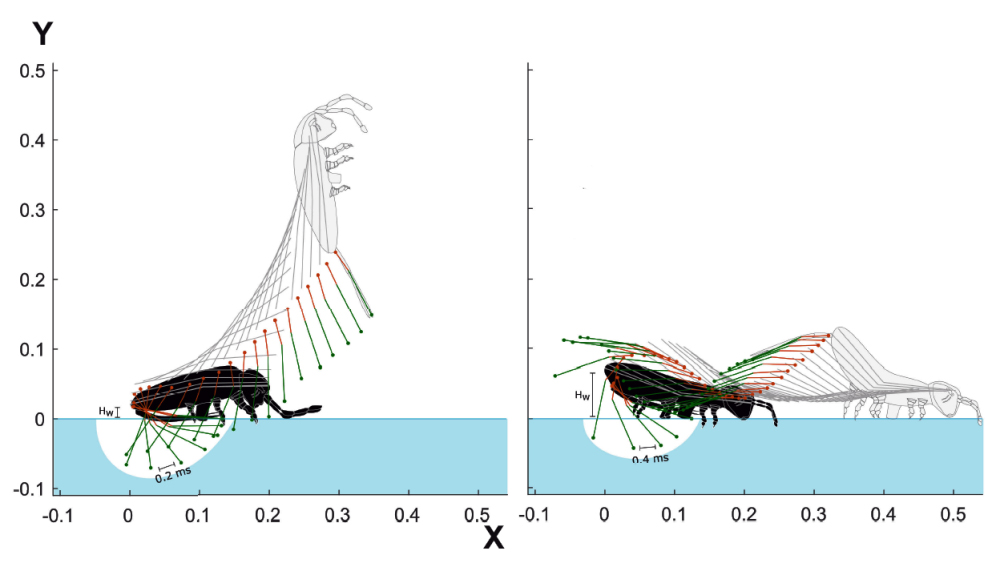
En complément de l’évocation des phases de décollage et d’atterrissage, penchons-nous maintenant sur la phase de vol, au cours de laquelle le collembole adopte, trois postures caractéristiques.
Outre ces trois postures, évoquées ci-aprés, on constate, lors de la visualisation des vidéos, que le collembole effectue un grand nombre de mouvement comparable à des contorsions qui montrent que ce dernier opère, tout au long de sa trajectoire, des ajustements de faible amplitude qui pourraient avoir pour objet de corriger cette dernière ou de parfaire son positionnement en vue de la réception.
Sur le schéma ci-dessous, les trajectoires des collemboles sont repérées par des couleurs :
- Trajectoire de la tête en rouge.
- Trajectoire de l’extrémité abdominale en bleu.
- Trajectoire du point central (centre de gravité approximatif) en noir.
Description :
- En phase ascensionnelle, la furca demeure tendue et le collembole effectue un retournement arrière sur lui-même (sens inverse des aiguilles d'une montre).
- Durant le vol, elle se replie contre l’abdomen et le collembole peut effectuer alors un nombre variable de rotations
- À l’amorce de la phase descendante, le collembole adopte une posture arquée, propice à sa réception.
Durant la partie aérienne, sous la zone notée « alternance rouge-bleu », j’ai repéré deux portions de trajectoire [b1-b2] et [r1-r2]. On constate que ces dernières alternent leur position (dessus/dessous) par rapport à la trajectoire noire. Chaque alternance correspond à une rotation du collembole sur lui-même et permet de compter le nombre total des rotations effectuées durant le vol.
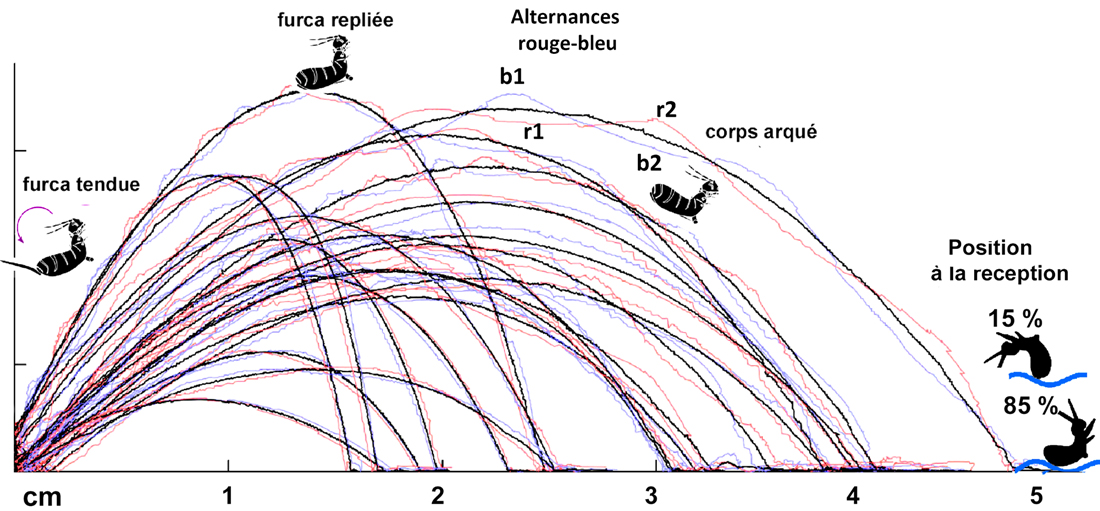
85% des collemboles, lors de leur arrivée, sont dans une posture arquée, ce qui permet au collophore d’être le point de premier contact avec l’eau. Ce mode de réception induit des atterrissages presque parfaits. 15 % des réceptions s’effectuent, en revanche, dans des positions ne permettant pas une stabilisation aussi rapide. Dans ces cas, les collemboles qui arrivent au contact de l’eau dans des postures moins appropriées se reçoivent de manière aléatoire sur diverses parties de leur corps. Le collophore ne peut alors jouer son rôle.
La stabilisation interviendra cependant à l’issue de mouvements de redressement effectuées généralement au cours du ou des rebonds. Les prises de vue révèlent alors des récupérations assez mouvementées, comme l’illustrent les figures ci-dessous.

Description :
- Les images (1, 2) montrent une réception en position inversée (sur le dos).
- Les antennes touchent l’eau en premier et le collembole s’écrase à la surface (3, 4).
- Il amorce d’un rebond (5).
- Retournement sur lui-même (6, 7).
- Réception sur la tête, mais en orientation ventrale (8).
- Fin du retournement permettant au collophore d’entrer au contact de l’eau (9).
- Le collophore joue son rôle de stabilisateur, aidé par les pattes arrières étendues (10).
Les phases d'atterrissage qui ont été chronométrées revèlent qu’une stabilisation réalisée à partir d’une mauvaise position de réception peut durer jusqu’à 10 fois plus longtemps qu’une réception de type ventrale avec usage immédiat du collophore, soit environ 44 ms.
Remarque : Les postures de vol décrites ci-aprés sont difficiles à restituer avec des images statiques.Je conseille de consulter les extraites vidéo que e mets à disposition par l'intermédiaire des liens suivants
Ci-dessous, quelques positions adoptées suivant les besoins au cours d’un vol :
 - Redressement en position arquée (1).
- Redressement en position arquée (1).
- Rotation d’amortissement en position arquée avec pattes tendues (2).
- Repli en forme de boule, pattes contre l'abdomen, pour accélérer la descente (3)
- Posture à l’envers d’un collembole mort (4)
- Posture aérodynamique avec furca tendue en début d’envol (5)
- Recours aux des pattes antérieures pour amorti à la réception (6)
Chez le collemboles disposant d’une morphologie globulaire, comme Bourletiella hortensis, la maîtrise des sauts semble moins établie. Une étude [5] réalisée par des chercheurs de l’université japonaise de Tohuku montre que ce dernier, effectue des sauts dont les profil distincts de ceux des Arthropléones.
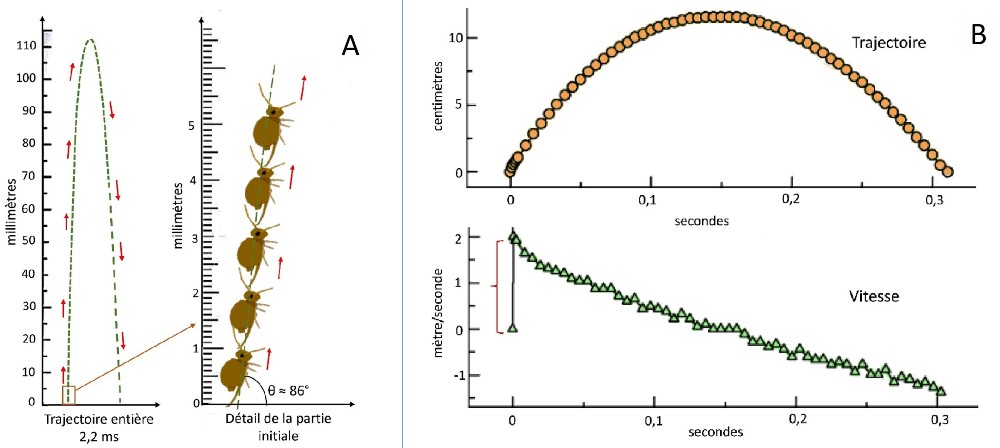
Les conditions de l’expérience diffèrent de celle précédemment décrite en raison de la nature du substrat. En effet, Bourletiella hortensis évolue sur des terrains solides qui exercent, sur les appuis de la furca, une incidence différente de celle d’un support liquide. Un autre déterminant important est la forme globulaire du collembole dont le profil aérodynamique se démarque sensiblement de celui d’un collembole au corps cylindrique.
Les chercheurs ont observé différents types de trajectoires qui montrent, pour le premier (fig.A ci-dessus), une phase ascensionnelle proches de la verticale (angle θ ≈ 86°) et une descente sensiblement symétrique.
La distance parcourue ici est faible. Elle permet au collembole de fuir un prédateur, mais ne peut être considérée véritablement comme un moyen de locomotion ordinaire. Ce collembole peut cependant effectuer des sauts plus longs, comme l’illustre la figure B, où la distance parcourue est égale à env. 100 fois la taille corporelle.
Ces valeurs doivent être modulées par le fait qu’un collembole globulaire effectue durant son vol jusqu’à 417 rotations/secondes. Ramené à la durée moyenne de vol (0.2 sec.), on obtient 83 rotations, soit environ 10 fois plus que celle observées généralement lors des sauts d’Arthropléones. La hauteur maximale atteinte est comparable à celle enregistrée sur la trajectoire précédente soit 11,5 cm.
La vitesse maximale de décollage atteint très rapidement 2 m/s, puis, elle décroit linéairement.
L’accélération atteint 300 m/s² en 1.65 ms., durant la phase repérée par une accolade rouge.
Accès aux vidéos :
Les differentes phases du vol de Isotomurus retardatus sont difficiles à traduire avec des montages photographiques. Ci-dessous accès à des extraits de vidéos d'où sont issus les montages de cet article.
Conclusion :
Le collembole semi-aquatique maîtrise en partie sa trajectoire. Ses mouvements durant le vol, lui permettent dans 85% des cas d'adopter la posture propre à une réception optimale.
Il serait utile de savoir dans quelle mesure, les autres postures qu’adopte le collembole durant le vol lui permettent d’agir sur des paramètres comme son Cx (coefficient de trainée) ou sa portance. On pourrait également étudier les comportements d’autres collemboles aux morphologies comparables évoluant sur différents substrats, pour chercher des similitudes ou des spécificités lors des vols.
Les études des sauts de Symphypléones révèlent par ailleurs des trajectoires moins maîtrisés et des réceptions plus aléatoires qui ne permettent pas d'arréter une conclusion.
Sources :
[1] : Victor M. Ortega-Jimenez a,b, Elio J. Challitaa,c, Baekgyeom Kimd, Hungtang Koc , Minseok Gwond, Je-Sung Kohd, and M. Saad Bhamlaa
aSchool of Chemical and Biomolecular Engineering, Georgia Institute of Technology, Atlanta, GA, USA; bSchool of Biology and Ecology, University of Maine, ME, USA;
cGeorge W. Woodruff School of Mechanical Engineering, Georgia Institute of Technology, Atlanta, GA, USA; dDept. Mechanical Engineering, Ajou University, Republic of Korea
[2] : Godrie Baptiste, « Rapports égalitaires dans la production des savoirs scientifiques. L’exemple des recherches participatives en santé mentale », Vie sociale, 2017/4 (n° 20), p. 99-116. DOI : 10.3917/vsoc.174.0099. URL : https://www.cairn.info/revue-vie-sociale-2017-4-page-99.htm
[3] : « On springtails (Hexapoda: Collembola): a morphofunctional study of the jumping apparatus »
Fábio Gonçalves de Lima Oliveira* - Frontiers in Zoology (2022)/ open acces.
[4] : Ce type de comparaison ne vaut que pour imager mon propos, car l’impact des accélérations sur des structures comme un corps humain, ne répondent pas, relativement à leur masse, à des valeurs linéaires car elles font intervenir des facteurs cubiques.