1) Introduction :
Dans l'article Évolution des collemboles, je souligne que ces derniers ont réussi à survivre à travers les ères géologiques, contrairement à d'autres espèces animales qui ont été victimes d'extinctions massives. Ces extinctions peuvent être liées à des changements biologiques dans l'environnement tels que des épisodes de prolifération végétale ou d'épuisement des ressources, ainsi qu'à des transformations physiques de parties plus ou moins étendues de la surface terrestre, provoquées par des éruptions volcaniques violentes, des éjections de lave et de cendres, ou encore des impacts d'astéroïdes.
Il convient alors de se demander comment les collemboles sont parvenus à faire face à de tels bouleversements de leur environnement. Des publications1-2 sur lesquelles je m’appuie ici ont retenu mon attention en apportant des éléments de réponse à cette question.
Deux options sont envisagées :
- La capacité des populations à se déplacer géographiquement à la recherche de conditions plus favorables.
- La capacité des populations à évoluer pour s'adapter à un environnement hostile.
2) Conditions de vie :
Les transformations des conditions de vie des espèces, au sein de leurs habitats originels, les condamnent à de nécessaires déplacements ou évolutions morphologiques et/ou physiologiques. Revoir l'article Déshydratation cryoprotectrice, dans lequel j’évoque un exemple d’adaptation de collemboles à des conditions extrêmes.
Les changements climatiques altèrent l'environnement de différentes manières, telles que des variations de température des eaux et de l'atmosphère, des fluctuations importantes du niveau des océans, des phases d'eutrophisation, des modifications de la composition chimique de l'atmosphère (teneurs en dioxyde de carbone, oxygène, méthane, etc.), des altérations de la photosynthèse et des épisodes glaciaires, pour n'en citer que quelques-uns. Il convient de noter que, même si ces évolutions et altérations passées se sont déroulées sur des périodes généralement brèves à l'échelle géologique, leurs effets se sont propagés à des rythmes relativement lents, sur une échelle temporelle allant de quelques centaines de milliers d'années à plusieurs millions d'années. En conséquence, ils ont probablement assez peu entravé l'évolution darwinienne des espèces.
3) Extinctions massives :
Contrairement aux évolutions « lentes » précitées, des événements accidentels, tels que les impacts d'astéroïdes, ont pu modifier l'environnement de manière plus fulgurante et, par conséquent, contrarier l'évolution naturelle des espèces. L'intensité de ces événements dépend de plusieurs facteurs pris en compte dans l'échelle de Turin. Cette dernière, qui comprend dix niveaux, intègre les probabilités de collisions, les tailles, les masses et les vitesses d'impact des astéroïdes et elle permet d'établir une projection de leurs conséquences. Il convient également de noter que l'IMCCE (Institut de mécanique céleste et de calcul des éphémérides) répertorie ces risques en fonction des astéroïdes identifiés à ce jour.
Les études géologiques et paléontologiques indiquent qu'en moyenne, tous les millénaires depuis la création de la Terre, un astéroïde de 100 mètres de diamètre frappe notre planète, causant la destruction de l'équivalent d'une grande ville ou provoquant un tsunami avec une vague de 35 mètres de haut qui inonde les zones côtières. Tous les 100 000 ans, un astéroïde de 500 mètres de diamètre détruit tout sur une surface comparable à celle de la France. Tous les 300 000 ans, un astéroïde d'environ 1000 mètres de diamètre détruit l'équivalent d'un grand pays, entraînant une perte estimée à 100 millions de vies humaines. Enfin, tous les 100 millions d'années, un astéroïde de 10 km provoque un cataclysme planétaire qui projette de la poussière dans l'atmosphère et crée un long hiver qui bloque la photosynthèse et perturbe la chaîne alimentaire, ce qui entraîne une période d'extinction massive.
Cependant, toutes les extinctions massives3 ne sont pas causées par des cataclysmes aussi violents. Seules deux d'entre elles sont attribuées à des collisions d'astéroïdes. Dans le tableau ci-dessous, les principales extinctions de masse connues sont répertoriées. Une ligne rouge indique l'apparition des premières traces fossilisées de coprolithes attribuées aux ancêtres des collemboles actuels.
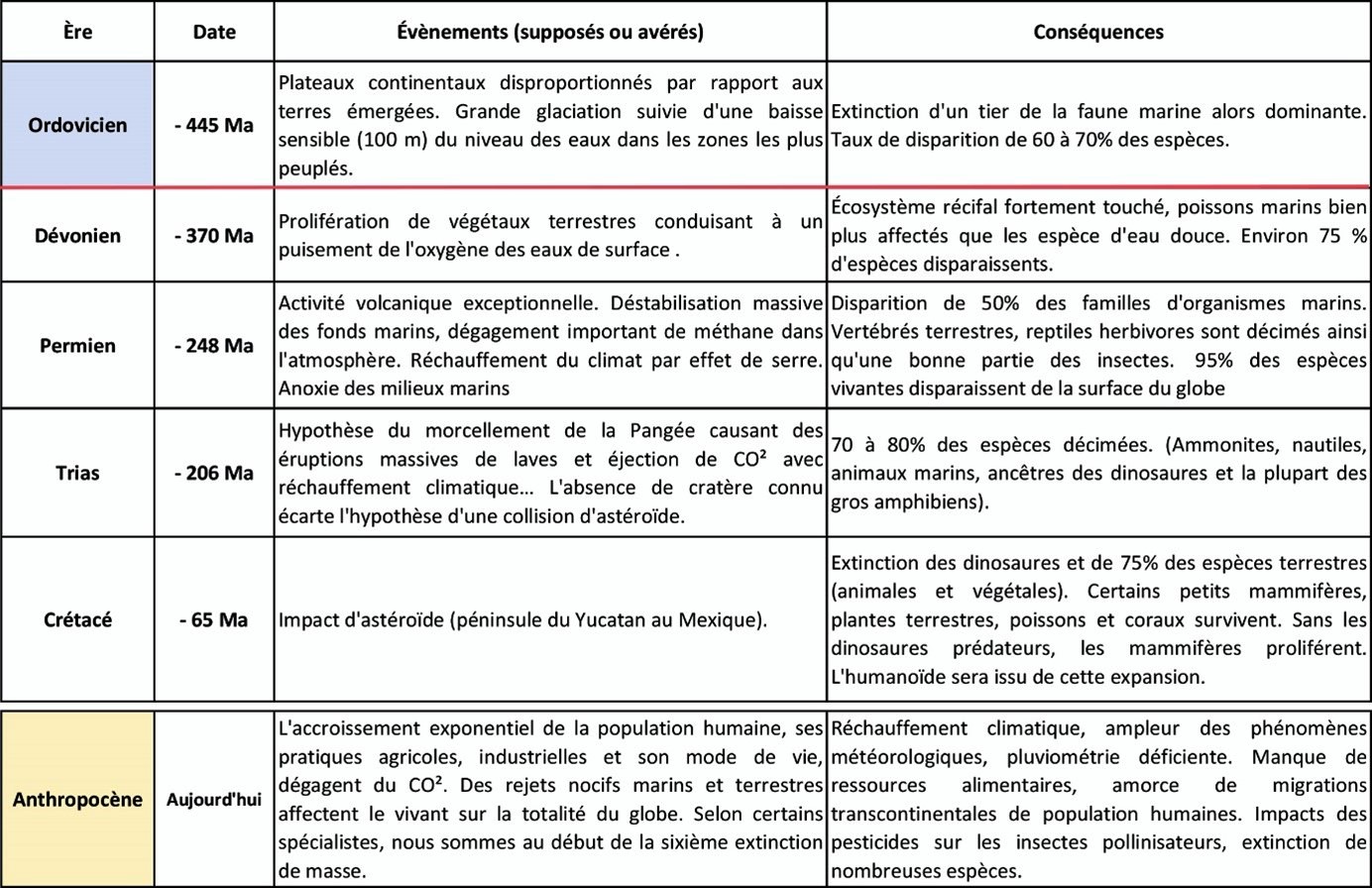
Les études menées sur les collemboles révèlent que ces arthropodes ont colonisé tous les biotopes de la Terre. Toutefois, lorsqu'on observe les communautés de collemboles au niveau des espèces, on constate des différences significatives entre les continents et les îles, en raison de leur isolement géographique, ainsi que des disparités à des échelles géographiques plus petites dues à des adaptations locales. Enfin, lorsqu'on observe des paysages modifiés par l'activité humaine, on constate que les collemboles les plus touchés sont ceux dont les déplacements sont les plus lents et les plus limités en distance.
C'est à ce stade que le déplacement ou l'adaptation offrent deux stratégies de survie possibles pour les collemboles. Cependant, la réponse à ces choix proposés n'est pas aussi tranchée que les termes "déplacement" et "adaptation" le laissent supposer. Les chercheurs apportent des nuances en notant que l'adaptation ne se traduit pas forcément sur le seul plan génétique et que le déplacement n'est pas l’unique moyen d'évitement.
Les collemboles sont des organismes capables de se déplacer pour faire face à diverses situations relatives à leur alimentation, leur reproduction, leurs mues, la toxicité de leur environnement et les risques de dessication. Cependant, leur capacité à se déplacer varie selon leur habitat, soit dans le sol ou en surface. La communication olfactive par le biais des phéromones joue un rôle important dans leur parcours sur de courtes distances dans les zones occupées par leur propre espèce (revoir communication chimique des collemboles )
Les caractéristiques morphologiques des collemboles sont étroitement liées à leur habitat. Les espèces hypogées* ont généralement des pattes et des antennes plus courtes que celles qui évoluent en zone épigée**. Les espèces en zone épigée peuvent se déplacer sur de plus longues distances par leurs propres moyens, mais leurs déplacements sont limités à des périmètres ne dépassant pas 200 mètres autour de leur lieu de vie, ce qui est insuffisant pour préserver les populations subissant des changements de leur biotope à l'échelle régionale ou continentale.
Au-delà de ce seuil de distance, le collembole doit recourir à d’autres vecteurs. C’est le cas pour la phorésie qui est un mode de déplacement utilisé par des animaux (dits phorétiques) qui se laissent transporter par d’autres animaux (hôtes) sur des distances dépassant leur capacité propre de déplacement.
Une étude2 effectuées à partir d’insectes emprisonnés dans des ambres fossiles datés de 16 millions d’années montre, en effet, que des collemboles Symphypléones ont usé de ce mode de locomotion, par voie aérienne (ce qui n’exclue pas les recours aux voies aquatiques/maritimes). Cette étude analyse deux insectes volant retrouvés emprisonnés dans un morceau d’ambre, sur lesquels se sont fixés des collemboles.
Les hôtes, un termite ailé et une fourmi, on permit à ces derniers d’être transporté par voie aérienne sur de longues distances. Les termites et les fourmis qui sont des insectes sociaux étaient abondants durant le Miocène, puisqu’ils représentent plus d’un tiers des insectes retrouvés dans les ambres de cette période. Ces insectes ailés ont donc très certainement été des vecteurs majeurs.
Certains caractères morphologiques ont permis à des collemboles de s’accrocher à leurs hôtes. Les antennes apparaissant comme le principal moyen de fixation, en raison d’un organe de serrage dont disposent certaines espèces. L’appareil buccal a été également utilisé, dans une moindre mesure, pour l’attachement phorétique.
Les chercheurs ont identifié d’autres stratégies de migration de collemboles susceptibles de se laisser emporter par les vents, par d’autres animaux de plus grande taille se déplaçant avec leurs pattes, mais également en se laissant entrainer par des eaux courantes (rafting) ou pousser par des vents à la surface des eaux. Une limite apparait quant à la dispersion par les airs, peu appropriée sur de longues durées en raison de la sensibilité des collemboles à la dessication.
4) L’adaptation :
Les changements adaptatifs sont souvent mis en évidence par la comparaison des populations. Le séquençage de l'ADN permet de mieux comprendre les processus évolutifs en cours, revoir « Étude du code-barre ADN des collemboles ».
Des scientifiques ont découvert qu’une mouche drosophile était capable de s'adapter à la toxicité des métaux lourds en dupliquant le gène codant pour une protéine appelée "metallothiénine". Cette protéine possède une haute affinité pour les ions métalliques et contribue à la détoxication des organes et des cellules contaminés.
À ce jour, aucune étude n'a révélé d'exemple similaire chez les collemboles, bien que des évolutions génétiques aient été constatées chez Orchesella cincta exposés à des stress métalliques, bactériens ou fongiques. Cependant, il est difficile de tirer des conclusions claires de ces observations. Des chercheurs ont également observé que chez des populations parthénogénétiques de Mesaphorura macrochaeta soumises à des stress, sont réapparu des modes de reproduction sexuée, suggérant une sélection plus adaptée des allèles***.
Folsomia candida, une espèce parthénogénétique connue pour être lucifuge et qui se déplace sur de courtes distances en quête de nourriture dispose d’une grande plasticité phénotypique**** qui lui permet de résister à la sécheresse, au froid, à la pollution et à de fortes concentrations de CO². Elle peut rapidement coloniser un nouvel environnement à partir d’une seule femelle.
Des changements non adaptatifs peuvent résulter d'isolements géographiques, comme dans le cas des collemboles cavernicoles ou de certaines espèces qui vivent en Antarctique. Chez d'autres familles, la communication chimique permet aux femelles d'exercer une forme de sélection entre les spermatophores déposés par les mâles. Cela peut conduire à des comportements d'évitement et même à l'absence de reconnaissance, entraînant la scission de populations avec des caractéristiques génétiques spécifiques bien qu’elles aient la même origine.
En dehors des milieux hostiles ou instables, les collemboles peuvent s'adapter à des changements périodiques comme les saisonnalités ou des changements plus lents relatifs à la dynamique de la couverture végétale. Le déplacement sur de courtes distances permet alors de maintenir les populations dans un habitat favorable. On notera également que les changements de pratiques agricoles tels que le labour, l'irrigation, les traitements divers ou les modifications des cultures, peuvent brusquement modifier l'habitat d’une population de collemboles, qui n'est alors plus adaptée à son environnement.
Pour résumer, on peut dire que les stratégies d'adaptation des collemboles résultent de plusieurs facteurs, comme le montre le schéma ci-après, proposé par le chercheur Jean-François Ponge.
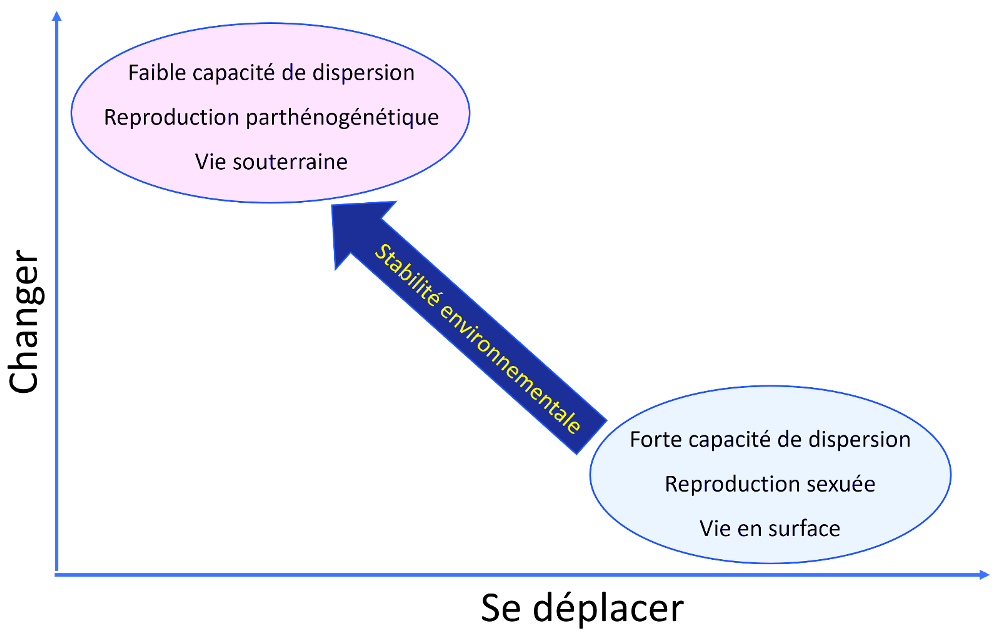
Les observations récentes mettent en évidence une flexibilité dans les réponses adaptatives des espèces face aux changements environnementaux. En effet, certaines espèces ayant une grande mobilité peuvent s'adapter aux changements en évitant de se déplacer, tandis que d'autres espèces qui ont une faible capacité de dispersion peuvent surmonter des distances supérieures à leur capacité de déplacement, grâce à des mécanismes encore mal connus.
 Les chercheurs ont cependant découvert que la protéine "métallothionéine" est surexprimée chez les Orchesella cincta qui occupent des zones contaminées par des métaux lourds (ci-contre, à gauche, Orchesella cincta, qui évolue en surface et dispose de pattes assez développées pour lui assurer une grande capacité de dispersion). Étonnamment, ce collembole n'a pas cherché à se déplacer pour fuir les zones contaminées...
Les chercheurs ont cependant découvert que la protéine "métallothionéine" est surexprimée chez les Orchesella cincta qui occupent des zones contaminées par des métaux lourds (ci-contre, à gauche, Orchesella cincta, qui évolue en surface et dispose de pattes assez développées pour lui assurer une grande capacité de dispersion). Étonnamment, ce collembole n'a pas cherché à se déplacer pour fuir les zones contaminées...
Par ailleurs, Folsomia candida et Mesaphorura (ci-dessous), deux espèces de collemboles qui vivent sous la surface du sol, ont été les premières à coloniser les zones minières à ciel ouvert, bien que leur faible capacité de déplacement aurait normalement empêché cette colonisation. Les chercheurs ont alors suggéré que ces collemboles ont pu être transportés sur de petits fragments de terre attachés aux pattes ou aux becs des oiseaux.

Ces observations montrent que les collemboles ont une grande plasticité phénotypique, leur permettant de s'adapter à des changements à petite échelle dans leur environnement, comme les perturbations naturelles ou agricoles. Ils peuvent également s'adapter verticalement ou survivre sous forme d'œufs en dormance. Cependant, il reste à savoir si les collemboles seront en mesure de faire face aux changements environnementaux à grande échelle, tels que le réchauffement climatique planétaire. Il est également important de se demander si les processus d'adaptation des collemboles atteindront leurs limites et conduiront à des extinctions.
5) Conclusion :
La recherche joue un rôle crucial dans la compréhension des réponses adaptatives des arthropodes aux changements environnementaux à grande échelle. Ces connaissances pourraient s'avérer utiles à pour l'humanité, qui semble avoir déjà un pied dans cette nouvelle ère Anthropocène au cours de laquelle elle sera inévitablement confrontée à la problématique de sa propre évolution. Dans ce cas "changer ou se déplacer" conserve toute sa pertinence....
Appendice :
* Espèces qui évoluent sous la surface du sol.
** Espèces qui évoluent à la surface des sols.
*** Les allèles sont des variantes d’un même gène.
**** La plasticité phénotypique se définit comme la capacité pour une cellule ou un individu, à partir d’une séquence génétique unique (appelé un génotype) présente dans son génome, d'exprimer une à plusieurs caractéristiques (appelées des phénotypes) selon différents contextes
Sources :
1 « Move or change, an eco-evolutionary dilemma: The case of Collembola » Jean-François Ponge Muséum National d’Histoire Naturelle, CNRS UMR 7179, 4 avenue du Petit Château, 91800, France. 7 February 2020 E-mail address: Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.. Pedobiologia - Journal of Soil Ecology 79 (2020) 150625 0031-4056/ © 2020 Elsevier
2 Ninon Robin, Cyrille D’haese, Phillip Barden. Fossil amber reveals springtails’ longstanding dispersal by social insects. BMC Evolutionary Biology, 2019, 19 (1), ff10.1186/s12862-019-1529-6ff. ffhal02378619f
3 Université de Laval – Canada - http://www2.ggl.ulaval.ca/personnel/bourque/s4/extinctions.masse.html
Images: P.G. sauf :
Orchesella cincta © Jérôme Picard - https://www.flickr.com/photos/jeromepi/ - Site internet: https://www.aquanat-photo.fr/